Схема переноса газов кровью




Перенос газов кровью
Кровь непрерывно переносит из легких в ткани кислород и из тканей в легкие углекислый газ. В артериальной крови, оттекающей от легких, кислорода содержится значительно больше, чем должно быть по физическим законам растворения газов в жидкостях. Это объясняется тем, что большая часть кислорода находится в крови не в растворенном, а в химически связанном состоянии. Кислород, поступающий из легочных альвеол в плазму крови, активно проникает в эритроциты и соединяется с гемоглобином, образуя непрочное химическое соединение — оксигемоглобин. Новые порции кислорода поступают из альвеол в плазму крови, а из нее в эритроциты до тех пор, пока почти весь гемоглобин не перейдет в оксигемоглобин. При дыхании атомсферным воздухом в обычных условиях 96% гемоглобина переходит в оксигемоглобин, и в результате в эритроцитах кислорода содержится в 60 раз больше, чем в плазме крови. Это и обеспечивает тканям необходимое для обмена количество кислорода.
Газообмен в тканях происходит по тому же принципу, что и в легких. При прохождении крови по кровеносным капиллярам различных органов кислород из области высокого парциального давления (плазма крови) переходит в область низкого парциального давления (тканевая жидкость). Из тканевой жидкости кислород поступает в клетки и сразу вступает в химические реакции окисления. Вследствие этого парциальное давление кислорода внутри клеток всегда равняется нулю. По мере выхода кислорода из плазмы крови оксигемоглобин переходит в гемоглобин, обеспечивая достаточную концентрацию кислорода в плазме. Превращению оксигемоглобина в гемоглобин способствуют многие факторы и, в частности, насыщение крови углекислым газом, и повышение температуры крови в органах (например, в мышцах во время их сокращения).
Углекислый газ, образующийся в клетках в процессе обмена, выходит в тканевую жидкость, создавая в ней высокое парциальное давление. В крови, протекающей по кровеносным капиллярам различных органов, парциальное давление углекислого газа значительно ниже, поэтому углекислый газ переходит из тканевой жидкости в кровь. Кровь содержит значительно больше углекислого газа, чем это возможно вследствие растворения его в жидкости. Это определяется также тем, что углекислый газ находится не только в растворенном состоянии в плазме, но и вступает в химическое соединение с гемоглобином эритроцитов и с солями плазмы. При участии специального фермента углекислый газ сравнительно легко соединяется также с водой плазмы крови, образуя угольную кислоту, которая в легких вновь распадается на углекислый газ и воду. Этим обеспечивается возможность выноса всей углекислоты, образующейся в тканях. Кровь, отдавшая кислород и насыщенная углекислым газом, называется венозной.
Венозная кровь поступает в легкие, где и происходит легочное дыхание.
Источник статьи: http://anfiz.ru/books/item/f00/s00/z0000002/st072.shtml
Схема переноса газов кровью
После поступления свежего воздуха в альвеолы начинается следующий этап дыхательного процесса: диффузия кислорода из альвеол в кровь и диффузия двуокиси углерода в обратном направлении — из крови в альвеолы. Процесс диффузии представляет собой беспорядочное движение молекул, прокладывающих себе дорогу через дыхательную мембрану и жидкости во всех направлениях. Однако в физиологии дыхания нас интересуют не только основные механизмы диффузии, но и ее скорость, что представляет собой намного более сложную проблему и потребует более глубоких знаний в области физики диффузии и обмена газов.
Физические основы диффузии и парциальные давления газов
Все газы, представляющие интерес для физиологии дыхания, являются простыми молекулами, которые свободно перемещаются в смеси. Этот процесс называют диффузией. Это справедливо и для газов, растворенных в жидкостях и тканях тела.
Для процесса диффузии необходимо наличие источника энергии. Энергия производится кинетическим движением самих молекул. При температуре выше абсолютного нуля молекулы находятся в постоянном движении. Это значит, что свободные молекулы, не связанные с другими молекулами, двигаются линейно на высокой скорости до встречи с другими молекулами. После столкновения их движение получит новое направление — до следующего столкновения. Таким образом, молекулы находятся в быстром и случайном движении среди себе подобных.
а) Диффузия газа одном направлении. Влияние градиента концентрации. Если в емкости или в растворе концентрация одного газа в одной зоне высокая, а в другой — низкая (для облегчения понимания просим вас изучить рисунок ниже), то суммарная диффузия газа будет направлена от зоны с высокой концентрацией в зону с низкой концентрацией: на рисунке в зоне А находится больше молекул, способных двигаться в направлении зоны Б, чем молекул, которые могут переместиться в обратном направлении, поэтому диффузия в каждом из направлений пропорциональна концентрации молекул, что на рисунке демонстрирует длина стрелок.
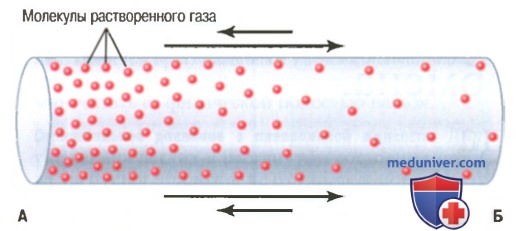 Диффузия кислорода из одной зоны (А) в другую (Б). Разница в длине стрелок представляет величину конечной диффузии
Диффузия кислорода из одной зоны (А) в другую (Б). Разница в длине стрелок представляет величину конечной диффузии
б) Давление газов в газовой смеси. Парциальные давления отдельных газов. Давление создается множественными ударами движущихся молекул о поверхность, поэтому давление газа на поверхности дыхательных ходов и альвеол пропорционально суммарной силе ударов о поверхность всех молекул данного газа в данный момент, т.е. давление газа прямо пропорционально концентрации молекул газа.
В физиологии дыхания мы имеем дело со смесями газов, состоящих главным образом из кислорода, азота и двуокиси углерода. Скорость диффузии каждого из них прямо пропорциональна давлению, создаваемому только этим газом, и это давление называют парциальным давлением данного газа. Далее приводим объяснение концепции парциального давления.
Воздух состоит примерно из 79% азота и 21% кислорода. Общее давление этой смеси на уровне моря равно 760 мм рт. ст. Из приведенного ранее объяснения молекулярных основ возникновения давления ясно, что доля каждого газа в давлении их смеси находится в прямой пропорции с его концентрацией, поэтому 79% из 760 мм рт. ст. давления воздуха создается азотом (600 мм рт. ст.) и 21% — кислородом (160 мм рт. ст.). Таким образом, парциальное давление азота в смеси составляет 600 мм рт. ст., парциальное давление кислорода — 160 мм рт.ст., а общее давление (760 мм рт. ст.) является суммой отдельных парциальных давлений. Парциальное давление отдельных газов обозначают PCO2, PO2, PN2, PH2O, PHe и т.д.
Источник статьи: http://meduniver.com/Medical/Physiology/862.html
Схема переноса газов кровью
Транспорт О 2 из атмосферы до периферических тканей организма представляет собой многоступенчатый процесс.
После того как воздух попадает в альвеолы легких, следующим этапом газообмена является диффузия кислорода из альвеол в кровь легочных капилляров и диффузия СО 2 из крови легочных капилляров в альвеолы. Диффузия представляет собой простое движение молекул через респираторную мембрану из области более высокого давления в область более низкого.
Таким образом, парциальное давление О 2 (PO 2 ) является одним из основных факторов, определяющих его транспорт в организме, причем не только в легких, но и во всем теле вплоть до периферических тканей.
Помимо градиента давления скорость диффузии определяется 1) растворимостью газа в жидкости; 2) площадью поверхности, через которую протекает диффузия; 3) расстоянием, которое газ должен пройти при диффузии; 4) молекулярным весом газа; 5) температурой жидкости. Поскольку в живом организме температура постоянна, она обычно не учитывается.
Применительно к альвеолокапиллярной мембране скорость диффузии газа будет зависеть от:
- толщины мембраны;
- площади поверхности мембраны;
- диффузионного коэффициента газа в мембране;
- градиента давления газа по обе стороны мембраны.
По мере транспорта О 2 от легких к периферическим тканям его парциальное давление снижается. Если в атмосферном воздухе при нормальном атмосферном давлении парциальное давление О 2 составляет 159 мм рт. ст., то в периферических тканях, в зависимости от уровня их обмена, — 35–85 мм рт. ст. Венозная кровь, поступающая в легкие, имеет РО 2 около 40 мм рт. ст.
Уже в альвеолярном воздухе содержание О 2 и его парциальное давление отличаются от атмосферного воздуха (13,6 % и 104 мм рт. ст. соответственно). Это происходит вследствие нескольких причин:
- альвеолярный воздух лишь частично замещается атмосферным во время каждого вдоха;
- О 2 постоянно абсорбируется из альвеолярного воздуха;
- СО 2 постоянно диффундирует из крови легочных капилляров в альвеолы;
- сухой атмосферный воздух, который поступает в дыхательные пути, увлажняется, насыщаясь водными парами, прежде чем достигает альвеол.
В артериальной крови, покидающей легкие, РО 2 уменьшается уже до 95 мм рт. ст. вследствие так называемого венозного примешивания. Дело в том, что определенное количество венозной крови (1–2 %) не аэрируется в легких, поступая напрямую в артериальное русло, что и приводит к некоторому снижению РО 2 . Увеличение объема шунтированной крови, которое происходит при гипоксии может приводить к существенному понижению артериального РО 2 .
Увеличение толщины альвеолокапиллярной мембраны, затрудняющее диффузию газов, часто является следствием отека — увеличения количества жидкости в межклеточном пространстве мембраны. Состояние характеризуется как интерстициальный отек легких. Кроме того, жидкость может накапливаться и в альвеолах, так что газам приходится проходить не только через мембрану, но и через жидкость, что характерно уже для альвеолярного отека легких. В большинстве случаев имеет место сочетание этих состояний той или иной степени выраженности. Имеет значение и исходное состояние легких. При ряде заболеваний, может возникнуть фиброз какого либо участка легких с утолщением альвеолокапиллярной мембраны. Поскольку скорость диффузии газов через мембрану обратно пропорциональна ее толщине, любой фактор, увеличивающий ее толщину более чем в два – три раза, может существенно нарушать нормальный газообмен.
Из капилляров кровеносного русла О 2 диффундирует в периферические ткани, поскольку парциальное давление О 2 в капиллярах выше, чем в тканях. РО 2 в интерстициальной жидкости вне капилляров составляет в среднем 40 мм рт. ст., тогда как в артериальной крови — 95 мм рт. ст. В венозной крови, покидающей капилляр, РО 2 также составляет около 40 мм рт. ст. Если скорость тканевого кровотока увеличивается, большие количества О 2 доставляются тканям и тканевое РО 2 увеличивается. Увеличение тканевого метаболизма приводит к снижению РО 2 как в периферических тканях, так и в венозной крови.
Поскольку О 2 постоянно используется тканями, внутриклеточное РО 2 всегда остается ниже интерстициального. Со снижением интерстициального РО 2 снижается и РО 2 внутри клеток, повышение интерстициального РО 2 ведет к повышению внутриклеточного РО 2 . Важным общим свойством живых организмов является большой запас функциональной прочности систем поддержания жизнедеятельности. Такая особенность характерна и для процессов газообмена. Для полного обеспечения процессов метаболизма достаточно РО 2 1–5 мм рт. ст., в то время как внутриклеточное РО 2 может варьировать от 5 до 60 мм рт. ст. (в зависимости от протяженности капиллярного русла). Поэтому организм человека и может функционировать при весьма существенном снижении РО 2 в атмосферном воздухе.
В нормальных условиях около 97 % кислорода от легких к тканям переносится в химически связанном виде гемоглобином. Лишь 3 % составляет О 2 растворенный в плазме крови. Следует учитывать, что способность гемоглобина связывать О 2 является ограниченной. Каждый грамм гемоглобина может максимально связать 1,34 мл О 2 . Эта так называемая константа Гюффнера. Соответственно кислородная емкость крови (т. е. максимальное общее количество кислорода, которое может быть перенесено кровью) будет находиться в прямой зависимости от содержания гемоглобина:
У здоровых людей с содержанием гемоглобина 150 г/л кислородная емкость крови составляет 201 мл О 2 /л крови. Реально переносимое количество мл О 2 обычно меньше.
Ключевым фактором, характеризующим количество кислорода, связанного с гемоглобином, является насыщение артериальной крови кислородом (сатурация, SaO 2 ). Оно выражает отношение между количеством кислорода, связанного с гемоглобином и кислородной емкостью крови:
Содержание кислорода в крови (контент, СаО 2 ) — это сумма связанного с гемоглобином и растворенного в плазме О 2 :
Очевидно, что СаО 2 зависит главным образом от фракции О 2 , связанной с гемоглобином.
Кровь содержит незначительное количество кислорода, не связанного с гемоглобином, а растворенного в плазме. Согласно закону Генри, количество растворенного кислорода пропорционально парциальному давлению О 2 и коэффициенту его растворимости, а растворимость О 2 в крови очень низка: только 0,0031 мл О 2 растворяется в 0,1 л крови при увеличении давления на 1 мм рт. ст. Таким образом, при РаО 2 равным 100 мм рт. ст., в 100 мл артериальной крови содержится только 0,31 мл растворенного О 2 . Со снижением РаО 2 количество растворенного в плазме О 2 станет еще меньше.
Содержание О 2 в связи с изменениями РаО 2 колеблется незначительно до тех пор, пока устойчиво поддерживается SaO 2 . Изменения содержания гемоглобина приводят к более заметным сдвигам СаО 2 . Нормальное СаО 2 равно 198 мл О 2 /л крови при условии, что PaO 2 = 100 мм рт. ст. , содержание гемоглобина 150 г/л, а SaO 2 = 97 % . Умеренная анемия (например гемоглобин 120 г/л) при поддержании нормального РаО 2 проявляется снижением СаО 2 до 160 мл О 2 /л крови.
Сродство гемоглобина к кислороду возрастает по мере последовательного связывания молекул О 2 , что придает кривой диссоциации оксигемоглобина сигмовидную или S-образную форму. Эта кривая, соотносящая изменения SaO 2 в зависимости от РаО 2 , важна для анализа процессов транспорта кислорода к периферическим тканям:
| Верхняя часть кривой (при РаО 2 > 60 мм рт. ст.) — относительно плоская. Это приводит к тому, что SaO 2 , а следовательно, и СаО 2 остаются достаточно постоянными, несмотря на значительные колебания РаО 2 . Повышение СаО 2 или транспорта кислорода в этой области кривой может быть достигнуто только за счет увеличения содержания гемоглобина (например, при переливании крови) или растворения в плазме крови кислорода (например при гипербарической оксигенации). Крутые средняя и нижняя часть кривой иллюстрируют то положение, что, хотя SaO 2 падает (когда РаО 2 оказывается ниже 60 мм рт. ст.), процесс насыщения гемоглобина кислородом продолжается, поскольку градиент РаО 2 между альвеолами и капиллярами сохраняется. Периферические ткани в этих условиях могут продолжать извлекать достаточное количество О 2 , несмотря на снижение капиллярного РО 2 . РаО 2 , при котором гемоглобин насыщен кислородом на 50 % (при 37 °С и рН 7,4) известно как Р 50 . Это общепринятая мера сродства гемоглобина к кислороду. Р 50 в крови человека в норме составляет 26,6 мм. рт. ст. Однако оно может изменяться при различных метаболических и физиологических условиях, воздействующих на процесс связывания кислорода гемоглобином. Когда сродство гемоглобина к кислороду падает, О 2 с большой легкостью переходит в ткани, и наоборот. Повышение Р 50 определяет сдвиг кривой диссоциации оксигемоглобина вправо. Сродство гемоглобина к кислороду снижается, указывая, что теперь требуется более высокое РаО 2 для поддержания SaO 2 на прежнем уровне. Более низкое сродство гемоглобина к кислороду означает повышенное высвобождение кислорода в тканях, но ухудшение связывания гемоглобина с О 2 в легких. Сдвиг кривой диссоциации влево и соответствующее снижение Р 50 указывает на повышенное сродство гемоглобина к кислороду — улучшение связывания в легких и ухудшение высвобождения О 2 в периферических тканях. На Р 50 и положение кривой диссоциации гемоглобина влияет несколько факторов, в частности, рН и температура. Для обеспечения обмена веществ в периферических тканях важное значение имеет не только общее содержание О 2 — ключевой физиологической переменной является его доставка. Доставка кислорода — это количество кислорода, транспортируемое к тканям в единицу времени. Чтобы поддержать аэробный метаболизм и предотвратить накопление лактата, периферические ткани должны постоянно снабжаться кислородом. Обстоятельства, которые определяют адекватность кислородного снабжения, разнообразны и включают состояние покоя, физическую нагрузку, гиперкатаболические состояния и инфекцию. Доставка кислорода к периферическим тканям зависит от количества кислорода в определенном объеме крови и уровня кровотока. Хотя объем кровотока к отдельным органам различен, в периферических тканях он в целом равен сердечному выбросу (CB). Системная доставка О 2 (DО 2 ) рассчитывается как DO 2 = СВ (л/мин) x CaO 2 (мл/л) . Нередко DО 2 рассчитывают с учетом площади тела. То есть на самом деле рассчитывают индекс DО 2 . Нормальными величинами индекса DО 2 считают 520–720 мл·мин -1 ·м -2 . Доставка кислорода падает при уменьшении сердечного выброса или снижении объемного содержания кислорода в артериальной крови. Это характерно для многих критических и терминальных состояний и чаще всего наблюдается при выраженных проявлениях сердечной недостаточности, повышении внутрилегочного шунтирования крови, централизации кровообращения, при различных пороках сердца, кардиохирургических и других длительных и травматичных вмешательствах. С практической точки зрения следует иметь в виду, что любое нарушение кровообращения будет ухудшать доставку О 2 . Потребление кислорода является заключительным этапом транспорта кислорода тканям и представляет собой кислородное обеспечение тканевого метаболизма. В условиях основного обмена взрослый человек потребляет около 250 мл О 2 в 1 мин. Однако скорость утилизации О 2 различными тканями значительно отличается. Потребление кислорода тканями (VO 2 ) — интегральный показатель, учитывающий как циркуляторный компонент транспорта кислорода (сердечный выброс), так и его гемический компонент (артериовенозное различие по кислороду, CaO 2 — CvO 2 ). Его можно определить по формуле: VO 2 = СВ x (CaO 2 — CvO 2 ) x 10 . Так же как и DО 2 , VO 2 часто выражают в виде индекса, то есть в перерасчете на площадь тела. Нормальными величинами индекса потребления кислорода являются 110–160 мл·мин -1 ·м -2 . В зависимости от состояния организма (покой, нагрузка или заболевание) меняется фракционное распределение СВ к органам. Более того, и экстракция кислорода тканями различных органов неодинакова. Например, миокард получает лишь малую фракцию СВ, но извлекает почти весь доставляемый кислород. Нормальный компенсаторный ответ на снижение кровотока проявляется в виде увеличения поглощения кислорода, достаточного для поддержания VO 2 на нормальном уровне. Падение сердечного выброса компенсируется увеличением артериовенозного различия по кислороду, и VO 2 остается неизменным. Снижение насыщения венозной крови кислородом отражает увеличение экстракции кислорода. Способность компенсировать снижение кровотока повышением поглощения кислорода является характерной особенностью микроциркуляторного русла практически всех органов и тканей за исключением сердца и диафрагмы. В них высокая экстракция кислорода из капиллярного ложа происходит уже в норме. Поэтому уровень кислорода в тканях сердца и диафрагмы весьма чувствителен даже к незначительным изменениям кровотока. Таким образом, основными факторами, определяющими доставку О 2 , являются:
Источник статьи: http://nmr.nioch.nsc.ru/naumenko/node3.html  Рено лагуна 1 — блок предохранителей Автомобиль Renault 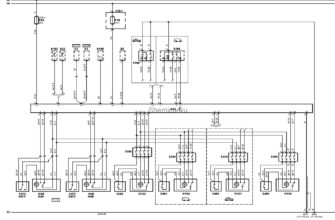 Схема автомобиля — Мерседес Спринтер Схемы электрооборудования 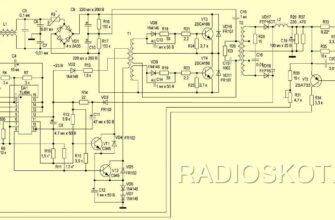 Схемы простых мощных зарядных устройств для аккумуляторов. |